

New observations on biology, ecology, host plants and parasitoids of four Larinus species are presented. Centaurea phrygia L. is confirmed as a host plant of Larinus obtusus Gyllenhal, 1835. The east Polish populations of Larinus obtusus were confirmed to be regionally monophagous on Centaurea stoebe L. and those of Larinus pollinis (Laicharting, 1781) on Carlina vulgaris L. Carduus crispus L. is identified for the first time as a host plant of Larinus sturnus (Schaller, 1783). Cirsium oleraceum (L.) Scop. and C. vulgare (Savi) Ten. were confirmed as host plants of Larinus sturnus, and Cirsium heterophyllum (L.) Hill as a host plant of Larinus turbinatus Gyllenhal, 1835. Life strategies including type of oviposition and development of these weevils are described and photographs are provided of immature stages, adults, host plants, biotopes and parasitoids.
Biology, host plant, Asteraceae, Cardueae, Carduus, Carlina, Centaurea, Cirsium, development, larva, pupa, Coleoptera, Curculionidae, Lixini, Larinus, Europe, Czech Republic, Poland, Romania
The weevil genus Larinus Dejean, 1821, belonging to the tribe Lixini Schoenherr, 1823, includes approximately 180 species [Csiki 1934][Ter-Minassian 1967][Gültekin 2006]. Approximately 140 species are known in the Palaearctic [Gültekin 2006], with 60 of these species being known in Europe [Mazur 2001][Alonso-Zarazaga 2009]. A further 40 species are recorded from the Ethiopian region, and only three from the Oriental region, four species were introduced to the Nearctic region [Smreczyński 1968][McClay 1988][Gültekin 2006], and one to New Zealand [Woodburn & Briese 1996][Gültekin 2006]. The genus Larinus is distinguished from other Lixinae by the following characters: a stocky body, rounded and slightly oblate dorso-ventrally, black in color, and sparsely hairy (Smreczyński 1968, Freude et al. 1983). The systematic position of this genus was assigned in the world Catalogue of Alonso-Zarazaga & Lyal [Alonso-Zarazaga & Lyal 1999] based on Csiki [Csiki 1934]. Recently, the genus Larinus has been divided in four subgenera: Larinus Dejean, 1821; Cryphopus Petri, 1907; Larinomesius Reitter, 1924 and Phyllonomeus Gistel, 1856 [Reitter 1924][Ter-Minassian 1967][Alonso-Zarazaga & Lyal 1999].
Larinus species have different habitat preferences; some of them are associated with xerothermic communities, whilst others inhabit wet meadows, pasture or ruderal communities [Koch 1992]. The studied species are generally monophagous or oligophagous, and they are associated with plants from the tribe Cardueae of the genera: Arctium L., Carduus L., Carlina L., Centaurea L., Cirsium Mill., Onopordum L. [Koch 1992]. The majority of Larinus species could be considered as potential biological control agents of weeds [Nikulina et al. 2004][Seastedt et al. 2007].
Larinus (Larinomesius) obtusus Gyllenhal, 1835 is known from Europe and the Near East region, with the northern border of its range in the central part of Poland [Wanat & Gosik 2003]. This species occurs in psammophilous grassland communities, dry meadows and ruderal communities [Koch 1992]. It is considered to be a stenotopic, thermophilous oligophage associated with the plant genus Centaurea [Koch 1992]. In 1991, it was introduced to the United States of America as a biological control agent for knapweeds [Seastedt et al. 2007].
Larinus (s. str.) pollinis (Laicharting, 1781) (= syn. L. brevis (Herbst, 1795)) is widely distributed from France across the southern and eastern regions of Europe, the Near East, Kazakhstan, and West Siberia to Korea [Mazur 2001]. L. pollinis is a stenotopic, xerothermic and oligophagous species that is associated with plants from the genus Carlina [Koch 1992][Mazur 2001].
Larinus (Phyllonomeus) sturnus (Schaller, 1783) is known from the southern part of Scandinavia, central and eastern Europe, North Africa and the Near East region (Burakowski et al. 1993). It is a stenotopic species associated with meadows and some other anthropogenic habitats. This oligophagous weevil develops on plant species from four genera: Arctium, Carduus, Centaurea and Cirsium [Zwölfer et al. 1971][Koch 1992].
Larinus (Phyllonomeus) turbinatus Gyllenhal, 1835 is distributed from the central part of Europe and North Africa to the central part of Asia [Burakowski et al. 1993][Alonso-Zarazaga 2009]. It is a stenotopic, thermophilous species associated with xerothermic, psammophilous grassland communities, pastures and ruderal vegetation [Koch 1992]. L. turbinatus is an oligophagous species that develops on plants from the genera Carduus and Cirsium [Koch 1992].
The present paper provides a detailed description of biology, host plants and ecology of four Larinus species. These data could be useful in the search for potential biological control agents of weeds (e.g. Carduus, Cirsium), and some weevils of this genus have already been successfully used for this purpose, e.g., Larinus carlinae (Olivier, 1807) [Seastedt et al. 2007].
The species were observed at the following localities (arranged according to host plants):
Larinus (Larinomesius) obtusus Gyllenhal, 1835
on wig knapweed (Centaurea phrygia L.) growing in a meadow under mountains: ROMANIA, prov. Alba, Rimetea, 33 km S of Cluj, brookside, N 46°28'57", E 23°34'64", 480 m a.s.l., 21.vii.2008, J. Skuhrovec lgt., coll. et det.;
on spotted knapweed (Centaurea stoebe L.) growing in psammophilous grassland communities (Corynephoro-Silenetum tataricae): POLAND centr. or., Wołczyny ad Włodawa [Fig. W63.8], UTM nets: FC80, 28.viii.2008; Stare Stulno ad Włodawa [Fig. W63.9], UTM nets: FC80, 28.viii.2008, all R. Gosik lgt., coll. et det.
Larinus (s. str.) pollinis (Laicharting, 1781)
on carline thistle (Carlina vulgaris L.): POLAND or., in xerothermic grassland communities (Brachypodio-Teucrietum) in Stawska Góra near Chełm, UTM nets: FB67, 13. and 20.viii.2009; sand grassland communities in Pieszowola near Urszulin, UTM nets: FC50, 10.ix.2009; POLAND mer. or., xerothermic grassland communities in Narol, UTM nets: FA68, 15.viii.2009, all R. Gosik lgt., coll. et det.
Larinus (Phyllonomeus) sturnus (Schaller, 1783)
on welted thistle (Carduus crispus L.) growing in psammophilous grassland communities: POLAND centr., sand grass communities in Stara Słupia, UTM nets: EB03, 12.vii.2009; sand grass communities in Pińczów, UTM nets: DA69, 12.vii.2009; R. Gosik lgt., coll. et det.;
on bull thistle (Cirsium vulgare (Savi) Ten.) growing in pastures: POLAND, Poleski National Park, Wola Wereszczyńska (FC50), 2. ix.2010, at a field road near to Lipniak (FC 40), Poleski National Park, 2.ix.2010; R. Gosik lgt., coll. et det.;
on cabbage thistle (Cirsium oleraceum (L.) Scop.) growing in wet meadows: POLAND centr. or., wet meadow (Cirsietum rivularis) in Łąki ad Nałęczów, UTM nets: EB88, 7. and 25.vi., 10. and 24.vi., 5. and 20.viii., 5.ix.2009; POLAND mer. or., wet meadow (Cirsietum rivularis) in Susiec, UTM nets: FA58, 20.viii.2009, R. Gosik lgt., coll. et det.
Larinus (Phyllonomeus) turbinatus Gyllenhal, 1835
on melancholy thistle (Cirsium heterophyllum L.) growing on a wet meadow (Filipendulion): CZECH REPUBLIC bor., Liberec env., Janov nad Nisou – Hrabětice (5257) [Fig. W63.19], N50°46'40.5'', E 15°41'8.09'', 740 m a.s.l., 8.vii.2008, J. Skuhrovec lgt., coll. et det.
The development of L. obtusus is known on eight Centaurea species [Tab. W63.20] [Zwölfer et al. 1971][Groppe 1992][Burakowski et al. 1993][Campobasso et al. 1999]. We observed its development on C. phrygia in NW Romania. The report of the development on C. stoebe in Poland is unique, because the described Polish population of L. obtusus is monophagous on C. stoebe.
In July 2008, the 5th International Meeting of the Curculio-Institute took place in NW-Romania (Transylvania). One of the localities visited was situated at a brookside in Rimetea (Alba province). The exact location is given under “Material and Methods”. The mid-mountain meadows were in full bloom, with flowers including Centaurea phrygia [Fig. W63.1].
The majority of knapweed plants were occupied by copulating pairs of L. obtusus [Tab. W63.21] [Fig. W63.2] or females which had already oviposited into the open flowers. Zwölfer & Stadler (2004) divided Larinus species into two groups according to their type of oviposition; (1) ‘Larinus turbinatus type’ – females lay eggs into the open flowerheads, and (2) ‘Larinus planus type’ – females lay eggs directly into the bud or, using their rostrum, create a hole at the bottom of a flowerhead in which to lay the eggs. Larinus obtusus [Fig. W63.2] belongs to the ‘Larinus turbinatus type’. We dissected several flowerheads in the field [Fig. W63.3] and a few of them were occupied by larvae or pupae of L. obtusus [Fig. W63.4] [Fig. W63.5] [Fig. W63.6].
One larva was parasitised by an unidentified hymenopteran parasitoid (Hymenoptera: Chalcidoidea) [Fig. W63.7]. The frequency of parasitism was very low.
Burakowski et al. [Burakowski et al. 1993] listed that the Polish populations of L. obtusus develop on only two Centaurea species; brown knapweed (C. jaceae L.) and yellow starthistle (C. solstitialis L.).
We observed adults and also immature stages exclusively on C. stoebe L. growing on psammophilous grassland communities (Corynephoro-Silenetum tataricae) [Fig. W63.8] [Fig. W63.9]. This location in Polesie Lowland Wołczyny in Poland also includes another possible host species, Centaurea scabiosa L., but the weevil developed only on C. stoebe [Fig. W63.10].
The first active adults of L. obtusus in the field were observed at the beginning of June. At the end of June, weevils copulated and females began to oviposit.
The first instar larvae were found in the flowerheads in the middle of July. Copulation and oviposition continued at this time, and females attacked new flowers. Mature larvae and pupae of L. obtusus [Fig. W63.11] [Fig. W63.12] were collected in the flowerheads from the end of July until the end of August. Larvae and pupae were only found on up to 5% of spotted knapweed specimens growing at the study site. We have never found more than one or two flowerheads per plant attacked by this weevil. We cannot with certainty state that females only ever lay one egg per flowerhead, but we have never found more than one larva or pupa in a single flowerhead. In contrast, the study by Burakowski et al. [Burakowski et al. 1993] reported the development of up to five larvae in one flowerhead. We doubt this because the flowerhead of this plant is small and we cannot conceive how five larvae could successfully complete their development inside one flowerhead of C. stoebe.
The immature stages of the weevil were only observed on a dozen plants of ca 500 exemplars examined at the sites investigated (ca 2.5%). Moreover, almost 75% of the immature stages were parasitized by an unidentified hymenopteran parasitoid.
The Polish populations of L. obtusus are extremely rare, stenotopic and regionally also monophagous, e.g. on C. stoebe [Fig. W63.10]. The effect of such a population on its host plant is really unpredictable, because “performance” was very low, only a few flowerheads were attacked. Therefore its value as a control agent would appear to be very low. However, the observed population occurs at the northern limit of its distribution. Probably, the less favourable environmental conditions for this species at its range edge make development problematic, resulting in low abundance and high mortality.
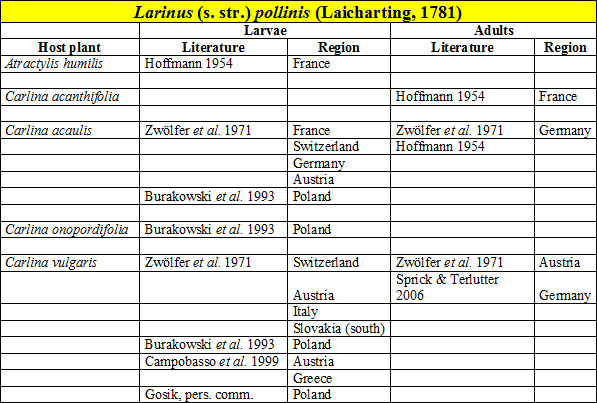
The development of L. pollinis is reliably known on only three Carlina species [Fig. W63.13] [Zwölfer et al. 1971][Burakowski et al. 1993][Campobasso et al. 1999]. Hoffmann [Hoffmann 1954] also reported Atractylis humilis L. and Carlina acanthifolia All. as its host plants. However, the data for Atractylis have not been confirmed, and it is suggested that this record is regarded with caution. Zwölfer et al. [Zwölfer et al. 1971] reported two bio-forms of L. pollinis.
The first active specimens of this xerothermophilous weevil were observed in the field attacking leaves in the middle of May. First instar larvae were found in the flowerheads in the middle of June. Mature larvae and pupae [Fig. W63.14] were collected from the beginning of August to the end of September. Some developmental stages of L. pollinis were observed in up to 60% of the carline thistle specimens growing at the studied locality. The flowerheads attacked by larvae exhibited decreased vitality. The flowerheads always contained only one larva. After their sclerotization and colouration, freshly hatched adults remained in the flowerheads even until the beginning of October, before overwintering in the soil nearby. We have never observed them overwintering in the flowerheads.
The proportion of parasitized larvae was low at this locality. Larvae from only 2 flowerheads from more than 50 flowerheads studied (4%) were attacked by an unidentified hymenopteran parasitoid.
Our observations correspond well with descriptions of L. pollinis by Cmoluch [Cmoluch 1965] and Burakowski et al. [Burakowski et al. 1993]. The development of L. pollinis in Poland was noted on silver thistle (Carlina acaulis L.), carline thistle (C. vulgaris L.) and C. onopordifolia Besser [Mazur 2001]. Cmoluch [Cmoluch 1965] reported its development in the Lublin Upland only on carline thistle (C. vulgaris). The formation of locally monophagous populations of oligo- or polyphagous species is well documented [Mazur 2001][Wanat 2004]. An explanation of this phenomenon could help us in the biological control by preventing its negative “off target” effects, however it has not yet been sufficiently clarified.
The development of L. sturnus is known on four different Cardueae genera, Arctium (3), Carduus (6), Cirsium (9) and Centaurea (3 species) [Tab. W63.22] [Zwölfer et al. 1971][Burakowski et al. 1993][Campobasso et al. 1999]. We studied its development on two Cirsium species (C. vulgare and C. oleraceum) and one Carduus species (C. crispus). The development on C. crispus observed in Poland is the first record for this host plant. Zwölfer et al. [Zwölfer et al. 1971] reported four bio-forms of L. sturnus (Arctium-, Carduus-, Centaurea- and “alpine” form).
In Poland, the northern limit of its range, L. sturnus is an expansive species. Recently, numerous populations were recorded from many new localities with adequate biotopes, e.g. wet meadows, pastures, ruderal communities, sand, xerothermic grassland communities and roadsides.
Copulation and oviposition (‘Larinus planus type’) [Tab. W63.23] were observed from the beginning of June until the end of July. Females laid eggs in a hole on the side of the bud. Different larval instars [Fig. W63.15] [Fig. W63.16] were found from the beginning of July until the end of August, and pupae [Fig. W63.17] from the middle of July until the end of September. The relatively long period of oviposition allowed us to study different larval instars and pupae in the field simultaneously. In C. oleraceum, immature stages were observed in up to 90% of the flowerheads [Fig. W63.18] at the study site. The larval abundance in C. crispus and C. vulgare was many times lower, ca 10%. We found only one or two flowerheads attacked by this weevil in one welted thistle (C. crispus); a similar abundance was also observed in the flowerheads of bull thistle (C. vulgare); on the contrary, on a single cabbage thistle plant (C. oleraceum), 6 flowerheads were attacked. It seems that attacked plants exhibit decreased vitality. In the laboratory (ca 20 °C, 13 : 11 L/D), pupation took 8 days, and sclerotization and colouring another 7 days. Afterwards, the young adults left the flowerheads.
The abundance of parasitized larvae was low at the locality. We found only three larvae parasited by an unidentified hymenopteran parasitoid.
Burakowski et al. [Burakowski et al. 1993] reported that the female always lays two eggs into each flowerhead, and both larvae developed simultaneously until one of them died because of their competition. We did not observe any similar behaviour and thus cannot confirm or refute this observation.
Koch [Koch 1992] presented L. sturnus as a stenotopic, hygrophilous weevil. These results did not correspond with our observations, or with Scherf [Scherf 1964] and Burakowski et al. [Burakowski et al. 1993], whose data on weevil’s life-cycle and biology correspond well with our studies. In our opinion, L. sturnus is a eurytopic species. It seems that the northern populations of this weevil are resistant and/or adapted to parasitoids. However L. sturnus seems to be a very useful control agent against selected weeds. Resolving its biotypes may prove essential.
The development of this species is reliably documented only on two closely related genera of Asteraceae, Carduus (2 species) and Cirsium (14 species) [Tab. W63.24] [Zwölfer et al. 1971][Burakowski et al. 1993][Campobasso et al. 1999]. However, adults of L. turbinatus were observed also on Silybum, Onopordum and Centaurea species [Tab. W63.24] [Zwölfer et al. 1971]. We studied its development on Cirsium heterophyllum in one locality in the Czech Republic [Fig. W63.19] near the border of Poland. Until recently, L. turbinatus was regarded as a rare species in Poland [Burakowski et al. 1993]. Recent observations suggested the expansion of L. turbinatus. Large populations were observed on xerothermic grassland communities (Cirsio-Brachypodion pinnati) and meadows (Arrhenatherion elatioris, Cirsietum rivularis) (Gosik 2006).
The first active adults were seen in the middle of June. Copulation and oviposition (‘Larinus turbinatus type’) [Tab. W63.25] were observed from the beginning of June until the middle of July. The different larval instars [Tab. W63.26] were found from the beginning of July until the beginning of August, and the pupae from the middle of July until the middle of August. The relatively long period of oviposition allowed us to study simultaneously the different larval instars and pupae in the field. No parasitized larvae were recorded at this locality. The adults were collected in Poland on Cirsium arvense (L.) Scop., occasionally on C. rivulare (Jacq.) All. and C. vulgare. Copulation of L. turbinatus on C. arvense and C. rivulare was observed in the first 10 days of June.
The adults were found in many different habitats, including wet meadows (e.g. development on C. heterophyllum in the Czech Republic). This indicates that its habitat preferences are wider than those suggested by Koch [Koch 1992].
Flowerheads of Cardueae plants contain a more varied, specific and well-known insect fauna than any other part of the plant. They represent a rich source of food, packed with achenes, and their inhabitants are protected from vertebrate predators by the tough spiny bracts [Redfern 1995]. The importance of pre-dispersal seed predators of thistles is well documented [Scherf 1964][Redfern 1995]. Weeds provide refugia and food for beneficial insects in agroecosystems [Batra 1982]. Biological control is the deliberate use of naturally occurring organisms to delimit the distribution and abundance of target weeds [Wilson & Randall 2003]. Increased interest in seed-feeding insects is based on their role in biological control. They may kill the weed directly (by weakening or stressing it) or indirectly (secondary infection with pathogens) (e.g. [Wilson and Randall 2003]). Seed-feeding insects play an important role in biological control [Redfern 1995]. Recently, several groups of pre-dispersal seed predators on thistles have been studied as potential biological control agents, e.g. Curculionidae [Louda & O’Brien 2002], and/or Tephritidae [Redfern 1995].
The accurate number of eggs deposited in a single flowerhead, the total number of flowerheads visited by one female, the accurate weevil abundance or the accurate parasitoids’ abundance were not determined. Exact results could only have been determined by detailed ecological studies. In the field, we recorded only baseline biological data. We always collected a few flowerheads for breeding [Fig. W63.3], and a few of them were immediately dissected in the field. Larvae and pupae were stored in ethylalcohol for future morphological studies [Gosik & Skuhrovec, submitted]. Moreover, detailed descriptions of the immature stages of Larinus species are important for further studies to promote the use of larvae of Larinus species as potential biological control agents against weeds (e.g. Carduus, Cirsium).
Although we did not estimate the weevils‘ abundance in flowerheads, their importance and usefulness as biological control agents for thistles [Louda et al. 2003] is suggested by their life strategies. Weevil larvae are able to destroy almost the whole receptacle and could prevent seeds from maturing, which will be tested in future studies [Skuhrovec et al. 2008]. Since there is little basic information on seed predation of all Cardueae species, there is still an urgent need to learn more about their seed predators.
We are indebted to Peter Sprick (Curculio Institute) and Levent Gültekin (Atatürk University, Faculty of Agriculture) for their valuable comments on the manuscript, to Max Barclay (Natural History Museum in London) for his kind linguistic help. The study was supported by a grant from the Ministry of Agriculture (Mze ČR) CZ0002700604, ME 10037 and the Foundation of Alexander von Humboldt (all JS).
Alonso-Zarazaga M.A. & Lyal, C.H. C. (1999): A World Catalogue of Families and Genera of Curculionoidea (Insecta: Coleoptera) (Excepting Scolytidae and Platypodidae). - S. C. P. Edition, Entomopraxis, Barcelona, 315 pp.
Alonso-Zarazaga M. A. (2009): Fauna Europaea, http://www.faunaeur.org.
Batra S.W.T. (1982): Biological Control of Weeds: Principles and Prospects, pp. 45-59. In Papavizas, G.C. (ed.): Biological Control in Crop Production, Beltsville Symposia in Agriculture Research. - Allenheld, Osmun and Co., Monclair, New Jersey and Granada Publ., Ltd., London.
Burakowski B., Mroczkowski, M. & Stefańska, J. (1993): Chrząszcze (Coleoptera) Ryjkowce Curculionidae. Katalog Fauny Polski, Warszawa, XXIII 19: 1-303.
Campobasso G., Colonnelli, E., Knutson, L., Terragitti, G. & Cristofaro, M. (1999): Wild plants and their associated insects in the Palearctic region, primarily Europe and the Middle East. - United States, Department of Agriculture, Agricultural Research Service, ARS-147, Washington, iv + 243 pp.
Cmoluch Z. (1965): Materialien Zur Kenntnis der Biologie und der geographischen Verbreitung von Larinus brevis Hbst. (Coleoptera, Curculionidae). - Annales UMCS, sec. C. 20 (2): 9-25.
Csiki E. (1934): Coleopterorum Catalogus auspiciis et auxilio W. Junk editus a S. Schenkling. Pars 134. Curculionidae: subfam. Cleoninae. - Junk, Berlin, 152 p.
Dieckmann L. (1983): Beiträge zur Insektenfauna der DDR: Coleoptera - Curculionidae (Tanymecinae, Leptopiinae, Cleoninae, Tanyrhynchinae, Cossoninae, Raymondionyminae, Bagoinae, Tanysphyrinae). - Beiträge zur Entomologie 33(2) 257-381.
Freude H., Harde, K. & Lohse, G. (1983): Die Käfer Mitteleuropas. Bd. 11. Rhynchophora II (Curculionidae II). - Goecke & Evers Verlag, Krefeld, 343 pp.
Gosik R. (2006): Weevils (Curculionoidea) of the middle part of the Bug River Valley. - Annales UMCS, sec. C. 61 (1): 7-69.
Gosik R. & Skuhrovec, J. (submitted): Descriptions of mature larvae of the genus Larinus (Coleoptera: Curculionidae, Lixinae). - Zootaxa.
Groppe, K. (1992): Larinus obtusus Gyll. (Col.: Curculionide). A candidate for biological control of diffuse and spotted knapweed. - C.A.B. Internat. Inst. Biol. Contr, Final Report, 46 pp.
Gültekin L. (2006): A new species of the weevil genus Larinus Dejean from Turkey and Syria (Coleoptera: Curculionidae: Lixinae). - Zootaxa 1248: 21-26.
Hoffmann A. (1954): Faune de France 59. Coléoptères Curculionidae. Deuxième Partie. - Paul Lechevalier, Paris, 487-1208 pp.
Koch K. (1992): Die Käfer Mitteleuropas. Ökologie. - Goecke & Evers, Krefeld, 371 pp.
Louda S.M., Arnett, A.E., Rand, T.A. & Russell, F.L. (2003): Invasiveness of Some Biological Control Insects and Adequacy of Their Ecological Risk Assessment and Regulation. - Conservation Biology 17: 73-82.
Louda S.M. & O’Brien, C.W. (2002): Unexpected Ecological Effects of Distributing the Exotic Weevil, Larinus planus (F.), for the Biological Control of Canada Thistle. - Conservation Biology 16: 717-727.
Mazur M. (2001): Ryjkowce kserotermiczne Polski (Coleoptera: Nemonychidae, Attelabidae, Apionidae, Curculionidae). - Studium zoogeograficzne. Monografie Fauny Polski, 22, 378 pp.
McClay A.S. (1988): The potential of Larinus planus (Coleoptera: Curculionidae), an accidentally introduced insect in North America, for biological control of Cirsium arvense (Compositae). - Proceedings of the VII International Symposium on Biological Control of Weeds. In E.S. Delfosse (ed.) in Instituto Sperimentale per la Patologia Vegetale Ministero dell’Agricoltura e delle Foreste 1990. pp.173-179.
Nikulina O., Gültekin, L. & Güçlü Ş. (2004): Larval morphology of the capitulum weevil, Larinus latus (Herbst) (Coleoptera, Curculionidae). - New Zealand Journal of Zoology 31: 23-26.
Redfern M. (1995): Insects and thistles. Naturalists’ Handbooks 4. - The Richmond Publishing Co. Ltd., Slough, England.
Reitter E. (1924): Die Larinus-Arten der Untergattungen Larinus s. str., Larinorhynchus, Larinomesius und Eustenopus aus Europa und den angrenzenden Gebieten. (Col. Curcul.). - Wiener Entomologische Zeitung 41(4/10): 61-77.
Scherf H. (1964): Die Entwicklungsstadien der mitteleuropäischen Curculioniden (Morphologie, Bionomie, Ökologie). - Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft 506: 336 pp.
Seastedt T.R., Knochel, D.G., Garmoe, M. & Shosky, S.A. (2007): Interactions and effects of multiple biological control insects on diffuse and spotted knapweed in the Front Range of Colorado. - Biological Control 42: 345-354.
Skuhrovec J. Koprdova, S. & Mikulka, J. (2008): How can seed feeders regulate dispersion of thistles, Cirsium arvense and C. heterophyllum? - Journal of Plant Diseases and Protection, Special Issue XXI: 281-284.
Smreczyński S. (1968): Podrodziny Tanymecinae, Cleoninae, Tanyrhynchinae, Hylobiinae. Klucze do oznaczania owadów Polski; Cz.XIX: Coleoptera. Zeszyt 98c: Curculionidae. - PWN. Warszawa, 106 pp.
Sprick P. & Terlutter, H. (2006): Funde bemerkenswerter phytophager Käfer in Westfalen (Schwerpunkt: Ostwestfalen) und angrenzenden Gebieten (Col., Nitiduloidea, Phalacridae, Cerambycidae, Chrysomelidae, Bruchidae et Curculionoidea) mit Anmerkungen zu aktuellen Ausbreitungsvorgängen und zum Status einiger Rüsselkäfer. - Mitteilungen der Arbeitsgemeinschaft westfälischer Entomologen 22(2): 33-83.
Ter-Minassian M.E. (1967): Zhuki-dolgonosiki podsemejstva Cleoninae fauny SSSR. Tsvetozhily i stebleedy (triba Lixini). - Nauka, Leningrad, 140 + 1 p. (English translation: Weevils of the Subfamily Cleoninae in the Fauna of the USSR. Tribe Lixini. USDA Agricultural Research Service, Washington, D. C. by Amerind Publishing Co. Pvt. Ltd., New Delhi, 1978. 166 pp.).
Volovnik S.V. (1994): On parasites and predators of Cleoninae weevils (Col., Curculionidae) in Ukrainian steppe. - Anz. Schädlingskde., Pflanzenschutz, Umweltschutz 67: 77-79.
Volovnik, S.V. (1995): (On distribution and ecology of some species of Cleoninae (Coleoptera, Curculionidae). III. Genus Larinus Germ.) - Entomologicheskoe obozrenie 74(2): 314-321 (in Russian).
Wanat M. (2004): Bagous elegans (Fabricius, 1801). In Głowaciński Z. & Nowacki J. (eds.) Polska Czerwona Księga Zwierząt - Bezkręgowce. 4 pp.
Wanat M. & Gosik, R. (2003): Materiały do znajomości ryjkowców (Insecta, Coleoptera: Curculionoidea) doliny Bugu. - Nowe Pam. Fizjograf., Warszawa 2(1-2): 31-52.
Wilson L.M. & Randall, C.B. (2003): Biology and Biological Control of Knapweed. - USDA-Forest Service FHTET-2001-07. 2nd Edition.
Woodburn T.L. & Briese, D.T. (1996): The contribution of biological control to the management of thistles. - Plant Protection 11(2): 250-253.
Zwölfer H., Frick, K.E. & Andres, L.A. (1971): A study of the host plant relationships of European members of the genus Larinus (Col.: Curculionidae). - Commonwealth Inst. Biol. Control, Techn. Bull. 13: 97-143.
Zwölfer H. & Stadler, B. (2004): The organization of pytophagous guilds in Cardueae flower heads: conclusions from null models. - Evolutionary Ecology Research 6: 1201-1218.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}