|
Weevil News |
No.10 |
4pp. |
15th March 2003 |
ISSN 1615-3472 |
|
|
Kozlowski,
M.W. (2003): Consumption of own eggs by curculionid females
(Coleoptera: Curculionidae: Curculioninae, Ceutorhynchinae) – Weevil News: http://www.curci.de,
No.10, 4pp., CURCULIO-Institut: Mönchengladbach (ISSN
1615-3472). |
|||||
by
Marek W. Kozlowski
(Figure 1) (Figure 2) (Figure 3) (Figure 4) (Clip 1) (Table 1)
Key words: Oviposition behavior, oophagy,
Coleoptera-Polyphaga, Curculionidae, Curculioninae, Ceutorhynchinae, Anthonomus, Cionus, Ceutorhynchus.
Females of five species
(Anthonomus rectirostris, Cionus
hortulanus, Ceutorhynchus napi, C. obstrictus, C. pallidactylus)
of the subfamilies Curculioninae and
Ceutorhynchinae were noticed consuming their own eggs that were released too
early or improperly during ovipositional process. The consumption of own eggs
in these rather strict phytophagous insects is probably an adaptative response
to either internal or external distortions of the ovipositional sequence resulting in improper egg deposition. It may
partially compensate for the loss of deficient nutrients incorporated to these
eggs. Own egg consumption can also form a prerequisite for the evolution toward
predatory or cannibalistic egg consumption in some weevils.
The Curculionidae or
true weevils is the family that comprises species living in majority as herbivores
on various plants (Balachowski, 1963; Smreczynski, 1972; Crowson, 1981;
Colonnelli et al., 1998). The subfamily Curculioninae sensu Crowson, (1955 ) followed by Smreczyński (1972) or
Kuschel (1995), is the most abundant among subfamilies of Curculionidae.
Species of Curculioninae live in tight and diverse relations with their
specific host plants. As larvae of Curculioninae are mainly plant
endoparasites, ovipositing females of Curculioninae use their prolonged rostral
beak to bore egg holes into specific host tissue. They deposit single eggs
(rarely several) at the bottom of these holes. Egg deposition is accomplished
by the aid of an ovitubus (retractable last three segments of the abdomen),
which females insert into the just bored hole. The action of egg deposition is
probably universal in females of this group and can be regarded as “rigid
moving pattern” (Tinbergen, 1989) generally composed from a chain of the
following movements: (1) egg hole formation and beak retracting, (2) turning
and localizing the entrance to the cavity with partially retracted ovitubus,
(3) hole penetration with the ovitubus and (4) egg deposition. More variable
among Curculioninae are the post-ovipositional events e.g. hole opening
covering with faeces (Rhynchaenus quercus
(L.), Apion frumentarium (L.), personal observation), adjacent tissue
manipulation [Anthonomus rubi
(Herbst) - Balachowski, (1963); personal observation].) or host organ marking
with deterring pheromone that prevents the same or the other females from repeatable
oviposition into the same organ (some Ceutorhynchus
Germar: Kozlowski et al., 1983,
Kozlowski, 1989 and Anthonomini spp. (unpublished observations on Anthonomus rectirostris (L.) and A. pomorum ). In five European
Curculionidae (two spp. of the subfamily Curculioninae - one of the
tribus Anthonomini and one of Cionini, and three spp. of the subfamily Ceutorhynchinae of the tribus
Ceutorhynchini), I sometimes observed
apparent distortions of normally rigid pre-ovipositional processes effected in early
egg release (“miscarrying”), usually aside the egg hole. Hence there have been no available data of this phenomenon, I give here a short description of such observations and
report that various Curculioninae females invariably devoured their “miscarried”
or imperfectly placed eggs, as soon as they released them from their oviducts.
Observations of early
release and consumption of own eggs were collected occasionally by laboratory
experimentation or documentation, originally aimed at investigations of mate
selection, oviposition, and pheromone marking of the oviposition site
(Kozlowski et al., 1983; Kozlowski, 1989; Kozlowski, 1991). In these
investigations, female weevils were placed separately on fresh fragments of
their host plants: Ceutorhynchus napi
Gyll. on topical part of a young rape plant (Brassica napus L.); C.
obstrictus Marsh. on young seedpods, C.
pallidactylus (Marsh.) on a rape plant leaf petiole with a leaf fragment,
and Anthonomus rectirostris on a
young bird cherry fruit (Prunus cerasus
L.) in various experimental arrangements (see Results). As early egg releases
happened irregularly and during both, pilot and experimental studies (less than
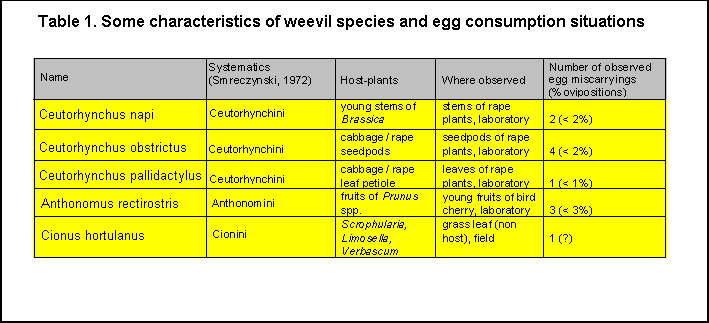
1-3 % of oviposition events), depending on a species (see Table 1) they were
originally recognized as “odd behaviors” and excluded from description and data
analyses. Consequently, no precise statistic of their frequency can be applied.
One case of egg consumption of the mullein weevil, Cionus hortulanus (Fourcroy, 1785), was documented in the field.
Pictures showing events of early egg release and consumption (Figure
1) were taken by a camera with flashlight (Ceutorhynchus napi) or extracted from
videotaping of weevil behaviour by the VHS and SVHS PAL laboratory unit (Ceutorhynchus spp. and Anthonomus rectirostris) as well as from
portable DVC PAL recording (Cionus hortulanus).
Improper release of eggs
by the weevil females were observed only during pre-ovipositional female
activities related with egg hole formation, turning and egg hole opening
localisation, except of C. hortulanus
where the behavioural context was not available. In each of the observed cases
of egg miscarrying, the females directly turned
to the miscarried eggs, chewed them with mandibles and apparently sipped out
their contents since the eggs became collapsed in seconds. Females then left
empty eggshells on the plant surface. Selected features of egg consumption
situations are summarized in (Table 1); picture documentation is presented on Figs 1-4 and Clip
1 (Clip 1).
There are following
comments to this process for particular species:
1. The
stem cabbage weevil, Ceutorhynchus napi.
Two cases of egg miscarrying and consumption were noticed in the
course of preliminary observation of oviposition behaviour and egg distribution
pattern (more than 100 oviposition acts; Kozlowski, unpublished). The cases of
egg miscarrying were noticed in two different females that had apparent
difficulties in hole opening localization with their ovipositors. In the
photographed act of egg miscarrying ((first case; (Figure 1)), the female
devoured the egg immediately after the picture was taken.
2. The
cabbage seedpod weevil, C. obstrictus.
All cases of egg consumption were noticed by the observations of females on
young rape seedpods by pilot and experimental investigations (more than 200
(ovipositions) of oviposition behaviour and seedpod marking in this species
(Kozlowski et al., 1983). In at least two cases of four noticed occurrences of
this behaviour, females that miscarried their eggs had also problems in
localization of freshly made oviposition holes.
3. The
less cabbage stem weevil, C.
pallidactylus. In this species females do not deposit single eggs but
several (3-9) ones into one egg hole (Guenthart, 1949). Ovipositing females
were observed either separately or with the presence of receptive males
(together more than 100 cases of ovipositions). In the documented single case of this egg consumption (Clip 1),
a male intensively courted the ovipositing female. This courtship as well as
aggressive attitude of female to the male (characteristic to this species -
Kozlowski, 1991) visibly distorted the co-ordination needed for egg hole
localisation. After consumption of the miscarried egg, the female preceded to
form another egg hole nearby.
4. The
fruit stone weevil, Anthonomus
rectirostris. Two cases of egg consumption were recorded in females
ovipositing (together, more than 100 ovipositions) during preliminary
observations and discrimination experiments related to post-ovipositional fruit
marking by the females into young fruits of the bird cherry. (Kozlowski,
unpublished; (Figure 2) and (Figure 3)).
5. The
mullein weevil, Cionus hortulanus. A
single case of this behaviour was observed in the field (Figure
4). A singular female staying on a grass leaf in the
vicinity of its host-plant Verbascum lychnitis L., densely populated by other
mullein weevils. In this case the female released the egg aside its host plant,
by unknown connection with the ovipositional sequence.
As the phenomenon of
early or imperfect egg deposition in Curculionidae seemed to happen rather
occasionally, it had not attracted attention of the students of weevil behavior
since no data on this event are available.
Defined reasons of the improper egg release are not known. It may have been
caused by internal (inaccurate neural pattern of turning movement) or external
(e.g. male presence in C. pallidactylus, uneven
structure of host plant in C. napi, artificial laboratory environments in
all but one cases, or internal distortions
of ovipositional co-ordination. Personal observations of rhythmical pulses of
the lateral part of the abdomen slightly before beak retraction suggests that
the action for egg release is triggered even before the female retracts its
beak and may cause egg miscarrying when the time of
hole localization is prolonged. The invariable consumptions of such eggs by the
females of 5 various species from subfamily Curculioninae suggest that this
phenomenon can be even more widespread among Curculioninae weevils and probably
also is present in plant-parasiting
weevils beyond this subfamily (the case of Larinus
sturnus observed by Knutelski, personal information). In all observed cases,
egg consumption can be considered as a special case of behavioral autophagy (e.
g. consumption of own body parts or products) being related neither to
predatory nor to aggressive and/or competitive acts aimed at other females.
Moreover, females consumed only those eggs that, being deposited incorrectly,
had apparently no chance of development (drying out, predators).
True weevils or
Curculionidae are considered as strict herbivorous, therefore consumption of
own eggs by weevil females can be a general exception from this rule. This
behaviour can be regarded as evidently adaptative. Egg production is expensive,
and miscarried ones, if consumed after release, can nutritionally contribute to
the production of the next eggs. On the other hand, innate inclination for the
consumption of own defective or defectively deposited eggs could trigger an
evolutionary route for cannibalistic or predatory egg consumption in these
weevils. In the surveyed literature
there is no available report on cannibalistic oophagy in weevils, but there are
two of predatory oophagy. The marked exception from general weevil herbivory is
South American Ludovix fasciatus Gyll.
Larvae of this species develop in egg batches of an Orthopterid that are found
by a mother female in the stems of the water hyacinth [Eichhornia crassipes (Mart.)]. It is interesting, that these eggs
are also important food for the adults of this weevil, that reach them by the aid of their prolonged
beaks. The contents are eaten during a long feeding bout (Klausnitzer, 1981).
The other example of egg predation characterizes species of the genus Archarius, that destroy and apparently
consume eggs of Pontania (L.) wasps
inside galls on willow leaves prior to deposition of their own eggs into galls
(personal observations, Kopelke, 1994). The same may happen in Curculio villosus (F.) where females
were observed with their beaks persistently “sounding” the galls of a gall wasp
Biorrhiza palliada Ol. (Kozlowski,
unpublished). One should not exclude an evolutionary scenario in which, in
selected species, the innate
predilections for the consumption of own, incorrectly deposited eggs have
redirected into the consumption of eggs of other insects to form new specific
predator-prey relationships.
Literature
Balachowski, A.E.
1963: Entomologie appliquée a l’agriculture. T. 1.
vol 2. Masson et Cie, Paris. 1391 pp.
Colonnelli, E., Louw, S., & Osella, G. (eds.)
1998: Taxonomy, ecology
and distribution of Curculionoidea (Coleoptera: Polyphaga). Proceedings of a Symposium in XX
International Congress of Entomology. Atti
Mus. Reg. Sci. Nat. Torino 6.: pp.
296.
Crowson, R. A. 1955: The natural
classification of the families of Coleoptera. Lloyd and Co. London
Crowson, R. A. 1981: The Biology of the
Coleoptera. Academic Press, London, xii
+801 pp.
Guenthart, E. 1949:
Beitraege zur Lebensweise und Bekämpfung von Ceutorhynchus quadridens Panz. und Ceutorhynchus napi Gyll. Mitteilungen Schweizerische Entomologische
Gesellschaft. 22: 779-780.
Klausnitzer, B. 1981: Wunderwelt der Käfer. Edition Leipzig. 211 pp.
Kopelke,
J.-P. 1994: Der Schmarotzerkomplex (Brutparasiten und Parasitoide) der
gallenbildenden Pontania-Arten
(Insecta: Hymenoptera: Tenthredinidae). Senckenbergiana Biologia 73: 83-133.
Kozlowski, M. K. 1989: Oviposition and host object marking by the
females of Ceutorhynchus floralis
(Coleoptera: Curculionidae): Entomologia Generalis 14: 197-201
Kozlowski, M.
K. 1991: Behavioural aspects of reproductive behaviour in Ceutorhynchus quadridens (Coleoptera:
Curculionidae) (in Polish): Publications of Warsaw Agricultural University, Wyd. SGGW, Warsaw, pp. 64
Kozlowski, M. K., S. Lux & J. Dmoch 1983: Oviposition
behaviour and pod marking in the cabbage seed weevil, Ceutorhynchus assimilis (Col.: Curculionidae). Entomologia
Experimentalis et Applicata 34:
277-282.
Kuschel, G. 1995: A phylogenetic
classification of Curculionidae to families and subfamilies. Mem. Entomological
Society of Washington 14: 5-33
Smreczynski, S. 1972: Klucze do oznaczania
owadow Polski. Cz. XIX, Zesz. 98 d, Coleoptera , Curculionidae). PWN, Warszawa, 195 pp.
Tinbergen, N. 1989: The study of instinct.
Oxford University Press. 256 pp.
Author:
Marek W.
Kozlowski
Department of Applied
Entomology, Warsaw Agricultural University SGGW
02-787 Warszawa, ul. Nowoursynowska, Poland
e-mail:
kozlowski@alpha.sggw.waw.pl
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}